



Le plan incliné





Copyright © Christian Delsanne pour CERB Club Des Éleveurs de Races Belges Ronquières Belgique
Le site du CERB a été créé en 1996.
Mises à jour en 2000, 2004, 2006, 2008 , 2010,2016 et 2021.









Club Des Éleveurs De Races Belges Ronquières
Informer | Inspirer
Club Des Éleveurs De Races Belges Ronquières
Informer | Inspirer

Il n’est pas nécessaire de s’inscrire pour naviguer librement sur ce site. Votre identité ne nous intéresse pas. La navigation est totalement anonyme


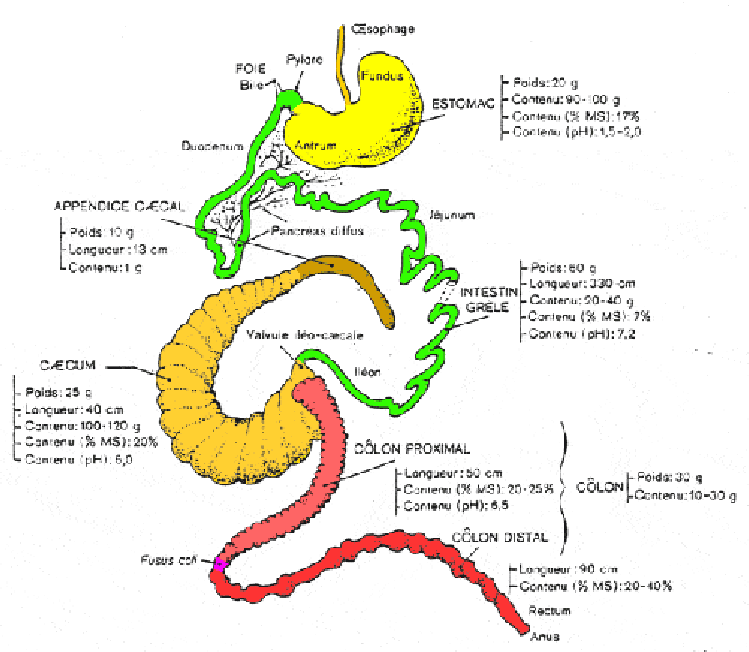
Situation chez l'adulte
Chez un lapin adulte (4-4,5 kg) ou subadulte (2,5 à 3 kg), le tube digestif a une longueur totale d'environ 4,5 à 5 mètres.
La bouche
Dans la bouche, les dents ont une croissance continue comme indiqué au chapitre 3. Leur rôle masticateur est très modéré. Les glandes salivaires (parotide, mandibulaire, sublinguale et zygomatique) produisent une salive contenant une faible quantité d'amylase (25 µMoles de maltose produites à partir le l'amidon, par mg de protéine salivaire, contre 250 à 450 pour le suc pancréatique). La concentration en amylase est d'ailleurs indépendante du taux d'amidon de la ration ou de l'état de jeûne de l'animal.
L'œsophage
L'œsophage est placé entre la trachée et la colonne vertébrale. Il ne permet de mouvement du bol alimentaire que dans la direction de l'estomac. Il n'y a jamais de reflux de l'estomac vers la bouche, même de manière accidentelle.
L'estomac
L'estomac est une poche allongée au revêtement muqueux. L'œsophage arrive dans l'estomac par le cardia. La partie "aveugle" de l'estomac correspond au fundus et la zone opposée est l'antrum qui se termine par le pylore .Ce dernier est muni d'un sphincter puissant qui régule les sorties d'aliment en direction de l'intestin grêle
La paroi secrète principalement de l'acide chlorhydrique, de la pepsine et des minéraux (Ca, K, Mg et Na). Le pH toujours fortement acide varie cependant assez sensiblement au cours de la journée, principalement dans la zone fundique (figure 12 ci-contre). Les pH les plus élevés sont observés en présence de cæcotrophes ( pH de 3,5) alors que le pH le plus fréquent est situé entre 1,5 et 2,0. La sécrétion de pepsine et d'électrolytes varie également principalement en fonction du rythme d'ingestion des caecotrophes. Le contenu de l'estomac représente 90 à 120g de matière fraîche chez un lapin de 9 semaines en fonction de l'heure de la journée. Sa teneur en matière sèche varie entre 16 et 21%.
Chez le très jeune lapereau (1ère semaine) la paroi stomacale secrète une pepsine dont le pH optimum est situé vers 1,8 - 2,4, ainsi que qu'une autre peptidase la rennine ou chymosine (pH optimum vers 3,4 - 3,8). A partir de 21 jours, le pH optimum de la pepsine s'abaisse aux environs de 1,2-1,8 tandis que la rennine n'est plus décelable chez le lapin de 45 ou 60 jours. Pendant la période d'allaitement, cette endopetidase est responsable de la coagulation du lait dans l'estomac par rupture de la chaîne de kappa-caséine. La sécrétion de pepsine ne devient quantitativement importante qu'à partir de l'âge de 30 jours environ. A l'inverse la lipase gastrique secrétée par une petite zone de la paroi stomacale autour du cardia a sa production maximum chez le lapereau de 30 jours, puis elle décroît rapidement entre 30 et 60 jours puis jusqu'à 180 jours. Elle n'est plus mesurable chez l'adulte.
L'intestin grêle
L'intestin grêle qui fait suite au pylore mesure environ 3 m de longueur pour un diamètre d'environ 0,8 à 1 centimètre. Il est classiquement divisé en duodénum, jéjunum et iléon, la partie terminale. Le canal cholédoque qui apporte la bile en provenance du foie débouche au début du duodénum, immédiatement après le pylore. Son ouverture dans le duodénum est régulée par le sphincter d'Oddi. Rappelons que chez le lapin la bile est sécrétée pratiquement en continu par le foie, puis stockée dans la vésicule biliaire avant son évacuation. Le canal pancréatique débouche vers la fin du duodénum à environ 40 cm du pylore. Sur la paroi, on observe de place en place des plaques de tissu lymphoïde d'environ 1 à 2 cm de diamètre. Il s'agit des plaques de Peyer. Les multiples glandes présentes dans la paroi de l'intestin grêle secrètent de nombreuses enzymes qui viennent compléter celles sécrétées par le pancréas.Le contenu de l'intestin grêle est liquide, surtout dans la première partie. En outre, il est normal de trouver des portions d'une dizaine de centimètres, vides de tout contenu. Le pH est légèrement alcalin dans la première partie (pH 7,2 - 7,5) et s'acidifie progressivement pour atteindre 6,2 - 6,5 à la fin de l'iléon.
Le cæcum
Cet intestin grêle débouche à la base du cæcum par le "sacculus rotondus" qui contient la valvule iléo-cæcal. Sa paroi est constituée de tissu lymphoïde. Le cæcum forme un second réservoir qui mesure environ 40-45 cm de longueur pour un diamètre moyen de 3 à 4 centimètres. Il contient 100 à 120 g d'une pâte homogène ayant une teneur en matière sèche (MS) de 22 pour cent en moyenne et un pH proche de 6 (figure 13 ci-après). La paroi du cæcum s'invagine selon une spirale qui fait 22 à 25 tours ou spires, augmentant ainsi la surface de muqueuse au contact du contenu cæcal. A son extrémité, l'appendice caecal (10-12 cm) a un diamètre nettement plus faible. Sa paroi est constituée de tissus lymphoïde.
Très près de l'abouchement de l'intestin grêle, c'est-à-dire de "l'entrée" du caecum, se trouve aussi le départ du côlon, autrement dit la "sortie". De ce fait, le caecum apparaît comme une impasse branchée en diverticule sur l'axe intestin-grêle-côlon .Les études de physiologie montrent cependant que cette impasse-réservoir est un lieu de passage obligé; le contenu circule de la base vers la pointe en passant au centre du caecum, puis revient vers la base, le long de la paroi.
Le colon
Après le caecum, on trouve un côlon d'environ 1,5 m. Il est d'abord caractérisé par la présence d'haustra (petits renflements en forme de poche) sur environ 50 cm : c'est le côlon proximal. Après une zone d'environ 1 à 1,5 cm portant les seuls muscles striés du tube digestif et appelée fusus coli, la paroi devient lisse dans sa partie terminale; cette partie est appelée côlon distal. Sa dernière partie est appelée rectum et se termine à l'anus. Ce dernier est porteur des glandes annales. Relativement plus développé chez le jeune que chez l'adulte, le tube digestif a pratiquement atteint sa taille définitive chez un lapin dès 2,5-2,7 kg, alors que l'animal ne pèse encore que 60-70 % au maximum de son poids adulte.Deux glandes importantes déversent leurs sécrétions dans l'intestin grêle : le foie et le pancréas. La bile, provenant du foie, contient des sels biliaires et de nombreuses substances organiques mais aucune enzyme: c'est une sécrétion qui aide à la digestion sans agir elle-même. A l'inverse, le suc pancréatique contient une quantité importante d'enzymes digestives permettant la dégradation des protéines (trypsine, chymotrypsine), de l'amidon (amylase) et des graisses (lipase).
Très globalement, il convient de retenir la longueur de l'intestin grêle (3-3,5 m) avec son faible contenu relatif, et l'importance des réservoirs que sont l'estomac et le caecum : 70 à 80 pour cent du contenu sec total du tube digestif sont en effet concentrés dans ces deux segments. Enfin, la teneur en eau du contenu peut varier très sensiblement d'un segment à l'autre, par suite des sécrétions de l'organisme ainsi que des absorptions d'eau.
Développement digestif en fonction de l'âge et du stade physiologique
Entre les âges de 3 et 5 semaines l'importance des segments antérieurs du tube digestif (estomac et intestin grêle) décroît tandis que celle des segments postérieurs s'accroît de manière significative (figure 14). Après l'âge de 7 semaines, l'ensemble des éléments du tube digestif a un développement moins rapide que celui du poids global du lapin, avec toutefois un "retard" de 2 semaines pour le côlon. Cas de la lapine reproductriceChez la lapine adulte, l'importance du tube digestif est réduite en fin de gestation, mais elle augmente fortement au cours de la lactation pour retrouver des valeurs relatives similaires à celles observée chez un jeune de 5 à 7 semaines (figure 14), ce qui démontre la plasticité du tube digestif.Compte tenu de l'évolution de la masse digestive et de la relative proportionnalité existant entre le poids de l'organe et celui de son contenu, le rendement à l'abattage s'améliore systématiquement avec l'âge, du moins chez les lapins de plus de 6 semaines (figures 15 et 16). En effet, le poids de la peau brute, l'autre élément important du rendement à l'abattage, ne varie que de 14,5 à 15,1% du poids vif entre 50 et 150 jours par exemple, alors que la masse digestive brute passe de près de 23% du poids vif à 50 jours à moins de 15% à 150 jours.
Physiologie de la digestion et caecotrophie
Les particules alimentaires consommées par le lapin arrivent rapidement dans l'estomac. Elles y trouvent un milieu très acide, y séjournent quelques heures (2 à 4 environ), mais y subissent peu de transformations chimiques. En fait, il y a une forte acidification entraînant la solubilisation de nombreuses substances, ainsi qu'un début d'hydrolyse des protéines sous l'action de la pepsine. Le contenu de l'estomac est progressivement "injecté" dans l'intestin grêle par petites salves grâce aux puissantes contractions stomacales. de l'estomac à l'intestin grêle
Dès l'entrée dans l'intestin grêle, le contenu est dilué par l'afflux de bile, par les premières sécrétions intestinales et enfin par le suc pancréatique. Sous l'action des enzymes contenues dans ces deux sécrétions, les éléments aisément dégradables sont libérés, franchissent la paroi de l'intestin et sont répartis par le sang en direction des cellules de l'organisme après le passage obligé par le foie (système porte). Les particules non dégradées, après un séjour total d'environ 1 heure 30 dans l'intestin grêle, entrent dans le caecum. Elles vont obligatoirement y séjourner un certain temps (de 2 à 12 heures). Pendant cette période elles subissent une attaque par les enzymes des bactéries vivant dans le caecum. Les éléments dégradables par cette nouvelle forme d'attaque sont libérés (acides gras volatils principalement) et à leur tour franchissent la paroi du tube digestif, puis sont repris par le sang. Le contenu du caecum à son tour est évacué vers le côlon. Il est constitué approximativement, pour moitié, par des particules alimentaires grosses et petites n'ayant pas été dégradées antérieurement et, pour l'autre moitié, par le corps des bactéries qui se sont développées dans le caecum aux dépens des éléments arrivant de l'intestin grêle et des restes de sécrétions digestives provenant de l'intestin grêle.
Jusqu'à ce stade, le fonctionnement du tube digestif du lapin n'est pas réellement différent de celui des autres monogastriques. Par contre, l'originalité est située dans le fonctionnement dualiste du côlon proximal. En effet, si le contenu caecal s'engage dans le côlon au cours du début de la matinée, il y subit peu de transformations biochimiques. La paroi colique sécrète un mucus qui enrobe progressivement les boules de contenu que les contractions de la paroi ont permis de former. Ces "boules" se trouvent réunies en grappes allongées. On les nomme crottes molles ou, plus savamment, "caecotrophes". Si, par contre, le contenu caecal s'engage dans le côlon à un autre moment dans la journée, son sort est différent. En effet, on observe alors dans le côlon proximal des successions de contractions de sens alterné, les unes tendant à évacuer "normalement" le contenu, les autres, à l'inverse, à le refouler vers le caecum. En raison des différences de puissance et de vitesse de déplacement de ces contractions, le contenu est en quelque sorte essoré comme une éponge que l'on presse. La fraction liquide, contenant les produits solubles et les petites particules (moins de 0,1 mm), est en grande partie refoulée vers le caecum, tandis que la fraction "solide", renfermant surtout les grosses particules (plus de 0,3 mm), forme les crottes dures qui seront évacuées dans les litières. En effet, grâce à ce fonctionnement dualiste, le côlon fabrique deux types de crottes: des crottes dures et des caecotrophes. Leur composition chimique est fournie au tableau 3. dualité d'excrétion et caecotrophie
Si les crottes dures sont évacuées dans les litières, à l'inverse, les caecotrophes sont récupérés par l'animal dès leur émission à l'anus. A cet effet, lors de l'émission, au cours d'une opération globale de toilettage, le lapin se retourne, aspire les crottes molles dès qu'elles sortent de l'anus, puis les avale sans les mâcher. De ce fait, le lapin peut, sans aucun inconvénient, pratiquer la récupération des caecotrophes même s'il est sur un sol grillagé. C'est pourquoi si un éleveur observe des cæcotrophes sous les cages de ses lapins, cela démontre que les animaux sont perturbés. En cas de litière accumulée, les crottes dures roulent les unes sur les autres lorsqu'elles arrivent au sol et forment ainsi des tas "étalés". Si des caecotrophes ne sont pas récupérés par l'animal en raison d'un stress temporaire ou permanent, le mucus qui les entoure tend à "coller" les crottes les unes aux autres. Dans ce cas, le tas de crottes sous les cages (à l'aplomb de l'arrière-train d'un lapin au moment où il consomme dans la mangeoire) est alors de forme "pointue".
Définition de la caecotrophie
Quand tout est normal, en fin de matinée, on retrouve les caecotrophes en grand nombre dans l'estomac où ils peuvent représenter jusqu'aux trois quarts du contenu. A partir de ce moment, le contenu des caecotrophes suit une digestion identique à celle des aliments "normaux". Compte tenu des fractions éventuellement recyclées une, deux voire 3 ou 4 fois, et de la nature des aliments, le transit digestif du lapin dure de 15 à 30 heures environ (20 heures en moyenne). Le fonctionnement général du tube digestif est schématisé sur la figure 17 ci dessous.
Valeur nutritive des caecotrophes
Il convient de rappeler que le contenu des caecotrophes est constitué pour moitié par des corps bactériens, et pour l'autre moitié par des résidus alimentaires non totalement dégradés, ainsi que par des restes des sécrétions du tube digestif. Les corps bactériens représentent un apport appréciable de protéines de bonne valeur biologique, ainsi que des vitamines hydrosolubles. La pratique de la caecotrophie présente donc, a priori, un intérêt nutritionnel non négligeable. Chez un lapin sain recevant un aliment complet équilibré, la pratique de la caecotrophie fournit à l'animal environ 15 à 25% des protéines ingérées quotidiennement et la totalité des vitamines B et C. Toutefois, si le mode de régulation et les quantités émises en limitent l'impact quantitatif vis à vis des protéines, l'apport est essentiel pour les vitamines hydrosolubles. De ce fait, un apport externe de vitamines hydrosolubles est souvent conseillé lorsqu'il existe un risque de perturbation digestive chez les lapins, comme c'est le cas pour les jours suivant le sevrage.
Le rôle du lest (des fibres)
Par contre, ce mode de fonctionnement particulier nécessite un apport de lest sous forme grossière. En effet, si l'aliment contient peu de particules grossières et/ou si celles-ci sont hautement digestibles, le refoulement vers le caecum fonctionne à son maximum et le contenu caecal s'appauvrit en éléments capables de nourrir les bactéries "normales" vivant dans le caecum. De ce fait, il apparaît un risque élevé de voir se développer des bactéries différentes dans ce milieu appauvri, une partie d'entre elles risquant d'être nocives. Il convient donc d'apporter, par voie alimentaire, un lest minimal qui permette aux animaux d'assurer un temps de séjour cæcal réduit et donc un transit digestif rapide. Classiquement, le lest alimentaire était appréhendé par la teneur en cellulose brute de l'aliment, mais ce mode d'expression reste peu précis. Aujourd'hui, les recommandations portent sur des apports minimums de ligno-cellulose (ADF) et de lignine. La figure 18 illustre le rôle de la quantité de fibres ingérée (composition fixe) sur les temps de séjour de l'aliment dans les différents segments du tube digestif. Il faut en retenir que les effets sur les temps de séjour dans l'estomac et dans l'intestin grêle tendent à se compenser, et les écarts entre les alimentations extrêmes ne dépassent pas 2 heures. Par contre, l'effet d'un apport réduit de fibres sur le temps de séjour du bol alimentaire dans le cæcum a une beaucoup plus grande amplitude, puisque ce temps passe de 9h 40 min. à 21h 30 min. avec l'apport le plus faible de fibres.
Rythme d'ingestion et rythme de caecotrophie sont liés
La régulation de la caecotrophie est dépendante de l'intégrité de la flore digestive et soumise au rythme d'ingestion. En effet, l'ingestion des caecotrophes est observée dans un délai de 8 à 12 heures, soit après le repas chez les lapins rationnés, soit après le pic d'ingestion chez les animaux nourris à volonté. Chez ces derniers, le rythme d'ingestion et, par voie de conséquence celui de la caecotrophie, est le résultat du rythme lumineux auquel ils sont soumis.Il faut signaler par ailleurs que la caecotrophie est également sous la dépendance de régulations internes encore mal connues. En particulier, l'ablation des glandes surrénales entraîne un arrêt de la pratique de la caecotrophie, et des injections de cortisone à ces animaux surrénalectomisés permettent de restituer un comportement normal. Ainsi, le transit digestif du lapin semble sous la dépendance étroite des sécrétions d'adrénaline. Une hypersécrétion associée à un stress entraîne un ralentissement de la motricité digestive et un risque élevé de troubles digestifs (mêmes conséquence qu'une carence en fibres alimentaires).Enfin, le comportement de caecotrophie apparaît chez le jeune lapin (domestique ou sauvage) aux environs de 3 semaines d'âge, au moment où les animaux commencent à consommer des aliments solides en plus du lait maternel.
| Nos expositions |
| Une exposition en bref |
| Marché paysan |
| Foire agricole de Soignies 2004 |
| Foire agricole de Soignies 2005 |
| Le souper annuel |
| Notre site |
| Présentation du club |
| Les canards |
| Les dindons |
| Les grandes volailles |
| Les volailles naines |
| Les lapins |
| Les pigeons |
| Les anatidés |
| Le canard domestique |
| Le canard sauvage "colvert" |
| Les canards de races belges |
| Liste |
| Histoire des dindons |
| Les dindons à Ronquières |
| Liste |
| Anatomie |
| Généralités |
| Liste des grandes volailles |
| L'aile |
| Bréchet |
| Pattes |
| Plumage |
| Queue |
| Squelette |
| Système circulatoire |
| Système digestif |
| Systeme musculaire |
| Système nerveux |
| Système respiratoire |
| Système uro génital |
| La tête |
| Origine |
| Définition |
| Domestication |
| Formation des races |
| Historique |
| Généralités |
| Liste des volailles naines |
| Photos de volailles naines |
| Barbus d'Anvers |
| Barbus d'Uccle |
| Brabançonne |
| Braekel |
| Fauve de hesbaye |
| Herve |
| Mehaigne |
| Tournaisis |
| Watermael |
| Généralités |
| Biologie du lapin |
| Liste des lapins |
| Photos de lapins |
| Argenté Belge |
| Beveren |
| Blanc de Termonde |
| Bleu de Ham |
| Gris perle de Hal |
| Steenkonijn |
| Generalites |
| Liste des pigeons |
| Photos de pigeons |
| Barbet Liegeois |
| Carneau |
| Cravaté gantois |
| Cravaté Liégeois |
| Culbutant belge |
| Haut volant belge |
| Ringslager belge |
| Smerle des flandres |
| Smijter |
| Speelderke |
| Voyageur belge |
| Voyageur Liégeois |
| L'agriculture |
| La fécondation |
| L'origine des oiseaux |
| En péril |
| La domestication |
| Le règne animal |
| Les fermes |
| Evolution |
| Système zoologique |
| Nomenclature |
| Distribution |
| L'adaptation |
| Communication |
| L'habitat |
| La vie sociale |
| Les expressions |
| Un jour un dicton |
| Contes et légendes |
| Fables de La Fontaine |
| A poil ! |
| Un canard boiteux |
| Du coq à l'âne |
| Poser un lapin |
| Fier comme un pou |
| Entre chien et loup |
| Sa langue au chat |
| Connu comme le loup blanc |
| C'est chouette ! |
| Janvier |
| Février |
| Mars |
| Avril |
| Mai |
| Juin |
| Juillet |
| Aout |
| Septembre |
| Octobre |
| Novembre |
| Décembre |