



Le plan incliné





Copyright © Christian Delsanne pour CERB Club Des Éleveurs de Races Belges Ronquières Belgique
Le site du CERB a été créé en 1996.
Mises à jour en 2000, 2004, 2006, 2008 , 2010,2016 et 2021.









Club Des Éleveurs De Races Belges Ronquières
Informer | Inspirer
Club Des Éleveurs De Races Belges Ronquières
Informer | Inspirer

Il n’est pas nécessaire de s’inscrire pour naviguer librement sur ce site. Votre identité ne nous intéresse pas. La navigation est totalement anonyme

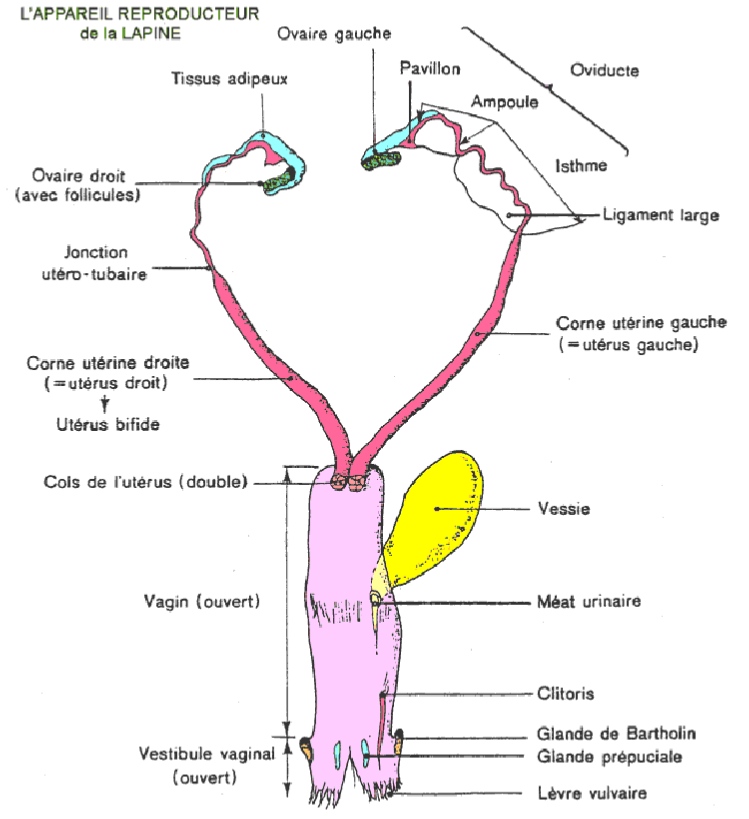
La position relative des différents organes est indiquée à la figure 34. Les ovaires sont ovoïdes ; ils atteignent 1 à 1,5 cm dans leur plus grande dimension. Sous chaque ovaire, le pavillon, l'ampoule et l'isthme constituent l'oviducte. Bien qu'extérieurement les cornes utérines soient réunies dans leur partie postérieure en un seul corps, il y a en réalité deux utérus indépendants de 7 cm environ, s'ouvrant séparément par deux conduits cervicaux dans le vagin qui est long de 6 à 10 centimètres. L'urètre s'ouvre dans la partie médiane du vagin au niveau du vestibule vaginal ; on peut distinguer les glandes de Bartholin et les glandes prépuciales. L'ensemble est soutenu par le ligament large qui a quatre points d'attache principaux sous la colonne vertébrale.
Comme pour le fœtus mâle, la différenciation sexuelle commence au 16e jour après la fécondation. Les divisions ovogoniales commencent le 21e jour de la vie fœtale et se poursuivent jusqu'à la naissance.
Après la naissance, les ovaires se développent nettement moins vite que l'ensemble du corps. Une accélération est observée à partir de 50-60 jours comme chez le jeune mâle, mais le ralentissement observé chez ce dernier après 110 jours n'est pas retrouvé chez la femelle. Les follicules primordiaux apparaissent dès le 13e jour après la naissance, les premiers follicules à antrum vers 65-70 jours .
Les femelles peuvent accepter pour la première fois l'accouplement vers 10 -12 semaines, mais à cet âge il n'entraîne pas encore l'ovulation. Par exemple sur une série expérimentale sur 80 lapines de 11 semaines présentées à un mâle adulte, 76% ont accepté de s'accoupler mais une seule a ovulé.
Compte tenu de l'absence de cycle œstrien et donc par d'œstrus spontané (voir plus loin), l'âge à la puberté est difficile à définir puisqu'il n'est pas possible de déterminer un âge au premier œstrus comme chez les autres espèces. L'âge à la puberté est donc déterminé par des critères indirects qui dépendent plus du type de population de lapines considéré que des individus eux-mêmes. Il dépend en particulier de la race. La précocité sexuelle est meilleure chez les races de petit ou moyen format (4 à 6 mois) que chez les races de grand format (5 à 8 mois). Dans les élevages commerciaux, les femelles sont couramment accouplées à 120-130 jours et montrent une bonne fertilité.
Du développement corporel.
La précocité est d'autant plus grande que la croissance a été rapide. Ainsi, des femelles alimentées à volonté sont pubères 3 semaines plus tôt que des femelles de même souche ne recevant chaque jour que 75 % du même aliment. Il est intéressant de constater que leur développement corporel est également retardé de 3 semaines.
La puberté des lapines est atteinte en général quand elles parviennent à 70-75 % du poids adulte. Cependant, il est souvent préférable d'attendre qu'elles aient atteint 80 % de ce poids pour les mettre en reproduction. Ces poids relatifs ne doivent cependant pas être considérés comme des seuils impératifs pour chaque individu, mais comme des limites valables pour la moyenne de la population. En effet, si le pourcentage de lapines capables d'ovuler s'accroît avec le poids vif moyen entre 14 et 20 semaines, à un âge donné il n'existe pas de différence de poids vif entre les lapines qui ovulent et celles qui n'ovulent pas .
En outre, comme indiqué plus haut, le comportement sexuel (acceptation de l'accouplement) apparaît bien avant l'aptitude à ovuler et à conduire une gestation. Ce comportement ne peut donc être utilisé par l'éleveur comme un signe de puberté, ce n'est qu'un signe précurseur. Seuls l'âge et le poids moyen de la population considérée doivent être pris en compte pour déterminer le moment de la puberté.
Oestrus et absence de cycle œstrien chez la lapine
Chez la plupart des mammifères domestiques, l'ovulation a lieu à intervalles réguliers au cours de la période des chaleurs, ou œstrus. L'intervalle entre deux périodes d'œstrus représente la durée du cycle oestrien (4 jours chez la rate, 17 jours chez la brebis, 21 jours chez la truie et la vache).
Par contre, la lapine ne présente pas de cycle œstrien avec apparition régulière des chaleurs au cours desquelles l'ovulation a lieu spontanément. Elle est considérée comme une femelle en œstrus plus ou moins permanent, et l'ovulation ne se produit que s'il y a eu accouplement. On considère donc qu'une femelle est en œstrus quand elle accepte de s'accoupler ; on la dit en diœstrus quand elle refuse. Pour ces deux états, on utilise aussi les termes de lapine réceptive ou non-réceptive.
De nombreuses observations montrent l'existence d'une alternance de périodes d'œstrus, pendant lesquelles la lapine accepte l'accouplement, et de périodes de diœstrus. Ces durées sont très variables d'un individu à l'autre, .Certaines lapines peuvent être en œstrus effectif pendant 28 jours consécutifs (ex.: lapine B), tandis que d'autres ne le sont que 2 jours en 4 semaines (ex.: lapine E). Actuellement, on ne sait pas prévoir les durées respectives des périodes d'œstrus et de diœstrus, ni quels sont les facteurs ambiants ou hormonaux qui les déterminent.
Toutefois, on constate que 90 % des femelles ayant la vulve rouge acceptent l'accouplement et ovulent. A l'inverse, 10 % seulement des femelles ayant une vulve blanche acceptent de s'accoupler et sont fécondées. La vulve rouge est donc une forte présomption d'œstrus, mais pas une preuve. Une lapine en œstrus caractérisé prend la position de lordose avec la croupe relevée, tandis qu'une lapine en diœstrus tend à se blottir dans un angle de cage ou à devenir agressive vis-à-vis du mâle.
Sur l'ovaire, les follicules à antrum qui n'ont pas pu évoluer jusqu'au stade ovulatoire (follicules de De Graaf) faute de stimulation (ni accouplement, ni administration d'hormone provoquant l'ovulation) régressent après 7 à 10 jours; ils sont plus ou moins rapidement remplacés par une nouvelle vague de follicules à antrum d'un diamètre supérieur à 800 µm. Ceux-ci restent à leur tour quelques jours sur l'ovaire au stade préovulatoire avant de régresser éventuellement à leur tour. Les cellules de la thèque entourant chaque follicules préovulatoires, secrètent des œstrogènes proportionnellement à leur masse. Le taux circulant de ces hormones n'est donc élevé que lorsqu'un nombre suffisant de follicules matures est présent sur l'ovaire. Cette information est intégrée par le système nerveux central qui modifie le comportement sexuel de la lapine et si le taux d'œstrogènes est "suffisamment" élevé la lapine devient réceptive "réceptive" à l'accouplement. Compte tenu de la variabilité entre individus, ce taux "suffisant" varie beaucoup d'une lapine à l'autre.
Le fait qu'une lapine ait des follicules au stade préovulatoire quasi en continu et une autre seulement quelques jours par mois, est une entrave à la synchronisation de la reproduction dans les élevages. L'administration à une lapine d'hormone folliculo-stimulante comme la PMSG, entraîne la maturation d'une nouvelle vague de follicules dans un délai de 48 à 72 heures, accompagnée d'une nette augmentation du taux d'œstrogènes et corrélativement l'apparition du comportement d'œstrus. Des résultats positifs sont régulièrement obtenus avec des injections de 8 à 25 UI de PMSG, en particulier chez la lapine allaitante, malgré l'effet inhibiteur partiel exercé par la prolactine sur l'activité ovarienne.
Des stimulations de l'œstrus peuvent aussi obtenues chez la lapine aussi par voie non hormonale, par la pratique des "biostimulations" faisant intervenir plus directement le système nerveux central et sa capacité d'intégration des facteurs environnant la lapine. Sur ce sujet le lecteur pourra se référer à la revue récente faite par Theau-Clément lors du congrès mondial de cuniculture qui s'est tenu à Valence (Espagne) en 2000. En quelques mots, ces biostimulations font appel à différents "stress" dont la séparation temporaire mère-jeunes chez les lapines allaitantes, à la modulation des rythmes lumineux ou à celle de l'alimentation.
Chez la plupart des mammifères, la progestérone sécrétée durant la gestation inhibe totalement l'œstrus, et la femelle en gestation refuse l'accouplement. Au contraire, la lapine gestante peut accepter l'accouplement tout au long de la gestation. Dans la deuxième moitié de la gestation, c'est même un comportement fréquent
De ce fait, l'éleveur ne peut compter sur le comportement sexuel des lapines pour savoir si elles sont ou non fécondées. Toutefois, une saillie éventuelle en cours de gestation n'a aucune conséquence néfaste pour les embryons portés par la femelle et ne provoque pas d'ovulation en raison de l'inhibition que la progestérone exerce au niveau central sur la libération de GnRH. Ainsi, contrairement à ce qui peut se produire chez la hase (femelle du lièvre), on n'observe jamais chez la lapine de phénomène de superfœtation (deux gestations simultanées à deux stades différents de développement). Par contre une injection de hCG ou de GnRH (1 à 10 µg/ kg P. vif) chez un lapine gestante peut entraîner une ovulation, ce qui risque de modifier le déroulement de la gestation en cours.
Dans un certain nombre d'espèces comme la truie, tout comportement d'œstrus est suspendu le temps de l'allaitement en raison du taux élevé de prolactine au cours de cette période. Chez la lapine, cette inhibition est loin d'être totale. Dans la majorité des cas, le taux de lapines réceptives (en œstrus spontané) diminue très significativement 4 à 5 jours après la mise bas pour remonter au dessus de 75% une dizaine de jours après le part. Le lien avec le taux de prolactine n'est cependant pas évident puisque les pics de prolactine enregistrés dans le sang après chaque tétée, ont une ampleur relativement stable de la mise bas au 25ème jour de lactation (74 ± 34 ng/ml) et ne diminuent qu'ensuite aux environs de 10 -15 ng/ml. Il faut également souligner que le taux de lapines en œstrus en fonction du délai écoulé depuis la mise bas varie beaucoup d'une expérience à l'autre.
L'ovulation
Normalement, l'ovulation est induite par les stimuli associés au coït ; elle a lieu 10 à 12 heures après la saillie. Dans la minute suivant l'accouplement, le taux d'ocytocine s'accroît tandis que celui de la prolactine décroît (figure 39). Cette décharge d'ocytocine semble avoir pour fonction de permettre aux spermatozoïdes de franchir les cols utérins et commencer à progresser dans l'utérus. Dans le même temps, l'hypothalamus envoie une décharge de GnRH qui atteint quasi immédiatement l'hypophyse par le système "porte" hypothalamo-hypophysaire. Seule une très faible fraction de cette décharge de GnRH se retrouve diluée dans le flot sanguin général, ce qui a pour conséquence que les taux circulants dans le sang périphérique n'ont aucune relation avec les taux physiologiques "efficaces".
Ensuite, en réponse à l'arrivée de GnRH, il y a une "décharge" de LH par l'anté-hypophyse. La concentration maximale est observée 90 minutes après le coït (figure 40). Une élévation beaucoup plus modeste du taux sanguin de FSH (l'autre gonadostimuline) est observée avec un maximum situé une demi heure plus tard. Cette décharge de LH permet l'évolution finale des gros follicules à antrum (diamètre supérieur à 0,8 mm) qui, en environ 10 heures, se transforment alors en follicule de De Graaf et libèrent chacun un ovule.
Compte tenu de cet enchaînement, on peut tenter de provoquer l'ovulation par des moyens artificiels en intervenant à différents niveaux. Une stimulation mécanique du vagin par action sur le cerveau peut provoquer des ovulations, mais les résultats sont très aléatoires (tableau 12). Par contre, des injections d'hormones GnRH, de hCG ou de LH donnent de bons résultats ; toutefois, des injections répétées de hCG ou LH entraînent une immunisation et une perte d'efficacité au-delà de la 4e ou de la 5e injection. Par contre, des injections répétées tous les 35 jours pendant 2 ans avec une GnRH de synthèse n'ont entraîné aucune baisse d'efficacité : 65 à 80 pour cent des lapines sont devenues gestantes avec l'injection suivie d'une insémination artificielle .
La fécondation et la gestation
Au moment de la rupture des follicules ovariens 10 à 11 heures après le coït, le pavillon de l'oviducte vient recouvrir l'ovaire. Dès leur libération, les ovocytes sont aspirés par le pavillon de l'oviducte et sont fécondables, mais ils ne seront fécondés qu'environ une heure et demie après leur émission.
Le sperme a été déposé par le mâle ou le dispositif d'insémination artificielle dans la partie supérieure du vagin à l'entrée des 2 cervix. La remontée des spermatozoïdes est rapide : ils peuvent atteindre le lieu de fécondation (dans la partie distale de l'ampoule, près de l'isthme) 30 minutes après le coït. Durant leur remontée, les spermatozoïdes effectuent une maturation qui les rend aptes à féconder les ovocytes. Sur les 150 à 200 millions de spermatozoïdes éjaculés, seulement 2 millions (1%) seront présents dans l'utérus ; ils rencontrent des obstacles principalement dans leur remontée au niveau du col utérin et de la jonction utéro-tubaire.
Au moment de la fécondation, sur chaque ovule une vingtaine de spermatozoïdes seulement sont présents, mais un seul traverse la membrane et assure la fécondation proprement dite.
L'œuf arrive dans l'utérus 72 heures après l'ovulation. Pendant la traversée de l'oviducte, l'œuf se divise. La paroi utérine se différencie, mais la dentelle utérine n'apparaîtra qu'entre 5 et 8 jours après le coït sous l'action de la progestérone. C'est la synchronisation de ces phénomènes qui permet l'implantation de l'œuf. L'implantation proprement dite s'effectue 7 jours après l'accouplement ; elle a lieu au stade blastocyte. La répartition des blastocytes est grossièrement équidistante dans chaque corne, mais il ne se produit pratiquement jamais que des blastocytes changent de corne utérine dans les conditions physiologiques normales. Du 3e au 12e jour suivant l'accouplement, le taux de progestérone ne cesse d'augmenter (multiplication par 4), puis reste relativement stationnaire pour diminuer rapidement dans les quelques jours précédant la mise bas)
Dans le même temps les taux d'œstrogènes subissent des modifications de moindre ampleur (variations de 1 à 2). Les pertes embryonnaires mesurées par comparaison du nombre de corps jaunes et du nombre d'embryons vivants sont en moyenne très importantes. En général, seulement 70 à 80 % des ovules pondus donnent finalement des lapereaux vivants à la naissance.
La majeure partie des mortalités embryonnaires se produit entre la fécondation (JO) et le 15e jour de la gestation (J15). La responsabilité de la mortalité embryonnaire incombe, d'une part, aux embryons (viabilité) et, d'autre part, à leur situation dans les cornes utérines. Mais certains facteurs extérieurs ont une influence, comme par exemple la saison et l'état physiologique des lapines (âge en particulier, ou état de lactation).
Par exemple chez la lapine simultanément allaitante et gestante post-partum (saillie féconde dans les 24 heures suivant une mise bas), la mortalité embryonnaire tardive est accrue par rapport celle observée chez une lapine seulement gestante dans les même conditions . Par contre une mortalité précoce importante est souvent en relation avec un taux faible de progestérone.
La pseudo gestation
Lorsque les ovules libérés ne sont pas fécondés, il se produit une pseudogestation qui dure 15 à 18 jours. Au début, le développement des corps jaunes et l'évolution de l'utérus sont les mêmes que pour une gestation, mais ils n'atteignent pas la taille ni le niveau de production de progestérone des corps jaunes gestatifs. Pendant toute cette période, la lapine n'est pas fécondable. Vers le 12e jour, ils commencent à régresser puis disparaissent par l'action d'un facteur lutéolytique sécrété par l'utérus, sous l'action de PGF2alpha. La fin de la pseudo gestation est accompagnée de l'apparition d'un comportement maternel et de la construction d'un nid liées à l'abaissement rapide du taux de progestérone sanguin.
Si la pseudo gestation est beaucoup utilisée dans les laboratoires de recherche sur la physiologie de la reproduction, elle est par contre très rare lorsque l'élevage est conduit en saillie naturelle. En effet, lorsqu'une femelle est saillie dans de mauvaises conditions, elle n'ovule pas, mais il est exceptionnel qu'à la suite d'une saillie naturelle on enregistre une ovulation sans aucune fécondation (cas d'un accouplement avec un mâle stérile, mais sexuellement actif). Par contre, les ovulations sans fécondation peuvent atteindre 20 à 30% des lapines inséminées artificiellement et ayant donc reçu une injection de GnRH pour les faire ovuler. Dans une telle situation, une injection de prostaglandine PGF2alpha effectuée au 10-11ème jour de la pseudogestation permet d'arrêter cette dernière et de féconder la lapine seulement 14 jours après une première insémination inféconde. Si on ne pratique pas ce traitement avec des prostaglandines, il est nécessaire d'attendre une semaine de plus pour tenter une nouvelle fécondation de la lapine. Généralement l'intervalle minimum respecté est de 3 semaines entre deux inséminations, donc entre 2 injections de GnRH.
Il existe cependant une autre situation particulière où les pseudo gestations peuvent être fréquentes si l'éleveur n'y prend pas garde: Cela concerne les lapines élevées en groupe. En effet, lorsque plusieurs lapines vivent dans une même cage, la femelle dominante chevauche les autres lapines. Dans ces conditions, les lapines dominées en œstrus peuvent ovuler et enclencher une pseudo gestation. C'est la raison pour laquelle il est toujours vivement conseillé de séparer les lapines futures reproductrices dans des cages individuelles au moins 3 semaines avant la date prévue pour leur première saillie ou insémination. Ce délai de 3 semaines garantit qu'une éventuelle pseudo gestation précoce est terminée et assure qu'aucune autre n'est enclenchée.
La mise bas
Le mécanisme de la parturition est assez mal connu. Il semble toutefois que le niveau de sécrétion des corticostéroïdes par les surrénales des jeunes lapereaux joue un rôle, comme c'est le cas dans d'autres espèces, pour donner le signal de la parturition. Les prostaglandines type PGF2a jouent également un rôle dans le déclenchement du part. A la fin de la gestation, la lapine construit un nid avec ses poils et la litière (paille, copeaux, etc.) mise à sa disposition. Les poils utilisés sont ceux de l'abdomen. En les retirant, la lapine dégage les tétines, ce qui en facilitera l'accès aux lapereaux Ce comportement est lié à une augmentation du rapport oestrogène/progestérone et à la sécrétion de prolactine. Parfois, la lapine ne construit pas le nid, ou elle met bas hors de la boîte à nid. Ce défaut comportemental est observé essentiellement lors de la première portée des lapines.
La mise bas dure de 10 à 20 minutes, sans relation très nette avec l'effectif de la portée. Quelques fois (au maximum 1 à 2% des mises bas) la lapine peut mettre bas en 2 fois espacées de plusieurs heures, il s'agit de situations exceptionnelles mais qu'il ne convient pas de considérer comme "pathologique". Le nombre de lapereaux par mise bas peut varier dans les cas extrêmes de 1 jusqu'à 20. Les portées les plus fréquemment rencontrées vont de 3 à 12 lapereaux ; les moyennes dans les élevages se situent entre 8 et 10 lapereaux par portée, mais cela reste très variable. Dans les 10 à 30 minutes suivant le début de la mise bas, la femelle a rapidement nettoyé les lapereaux des résidus d'enveloppes fœtales qui restaient sur leur corps. Dans le même temps le lapine consomme les placentas. L'observation de placenta dans la boite à nid plus d'une heure après la mise bas peut être considéré comme une anomalie.
Une gestation normale dure de 30 à 32 jours. Une mise bas après 29 jours de gestation correspond à la naissance de prématurés. Parfois le gestation est prolongée jusqu'à 33 ou 34 jours; dans ce cas il n'y a très généralement que 1 à 3 lapereaux, et souvent des mort-nés. Les lapereaux nés après 32 jours de gestation sont plus lourds au moment de leur naissance que ceux nés après une gestation de 30 jours seulement. En fait ils ont continué leur croissance in utero et pèsent à 32 jours de gestation pratiquement le même poids que des lapereaux de 2 jours nés après une gestation de 30 jours seulement. C'est une des raisons principales qui nous ont conduit à conseiller de considérer l'âge des lapereaux en prenant le moment de la saillie (ou de l'insémination) comme point de départ et non celui de la naissance.
Après la mise bas, l'utérus régresse très rapidement et perd plus de la moitié de son poids en moins de 48 heures. Comme déjà mentionné, la lapine est fécondable immédiatement après la mise bas et le sera tout au long de la période d'allaitement, avec des résultats cependant un peu moins "bons" pour les fécondations obtenues dans la semaine suivant la naissance des lapereaux.
Allaitement et lactation
La lactogénèse (synthèse du lait) est sous la dépendance de la prolactine. Pendant la gestation, elle est inhibée par les œstrogènes et la progestérone. A la parturition, il y a diminution rapide de la teneur en progestérone et, sous l'effet de la libération d'ocytocine, l'action de la prolactine est stimulée, ce qui permet la montée laiteuse dans une glande pré-développée. Ainsi au moment de la mise bas il y a séjà 50 à 80 g de lait dans les mamelles de la lapine. Ce type de lait est appelé colostrum. Il est consommé par les lapereaux au fur et à mesure des naissance : les premiers nés ont cliarement fini de téter quand "sortent" les derniers lapereaux de la portée malgré la bièveté de la mise bas (10 - 20 mn).
La libération du lait se produit de la façon suivante : lorsque la lapine vient donner à téter à sa portée, les stimuli créés par la tétée provoquent la sécrétion immédiate d'ocytocine, la pression intramammaire augmente, l'éjection du lait se produit et les lapereaux vident presque totalement la mamelle (80 à 90% du lait présent). Le taux d'ocytocine ne reste élevé que 3 à 5 minutes. La durée totale de la tétée (entrée-sortie de la boite à nid) n'est d'ailleurs que de 2 à 4 minutes. La concentration plasmatique d'ocytocine s'accroît de 40 pg/ml de plasma 2 jours après la mise bas, à 250 et 490 pg/ml au milieu puis en fin de lactation, sachant qu'un taux minimum de 20-25 pg/ml est nécessaire à l'enclenchement du processus d'éjection du lait.
A l'inverse, la durée de la tétée décroît lentement mais régulièrement avec l'avancée de la lactation : passage de 200 à 150 secondes par exemple entre le 14e et le 35e jour d'allaitement. Enfin cette durée est indépendante du nombre de lapereaux allaités et du fait qu'une lapine soit ou non simultanément gestante.
C'est la femelle qui fixe le rythme des tétées: une seule fois par 24 heures. La seule succion exercée par les lapereaux n'est pas suffisante pour déclencher la décharge d'ocytocine. Il faut la volonté de la mère. Dans quelques cas, la lapine peut donner à téter deux fois par 24 heures. Certains auteurs de l'équipe l'Université de Giessen (Allemagne) pensent avoir observé des allaitements jusqu'à 4 et 5 fois par jour, mais il semble bien que ces auteurs aient confondu des entrées répétées de la femelle dans la boite à nid avec des allaitements effectifs. En effet, ils n'ont pas (encore ?) été en mesure de prouver que du lait passe bien de la mère aux lapereaux lors des séjours répétés de la lapine dans sa boite à nid.
Une à cinq minutes après la fin de la tétée, on constate une décharge de prolactine (70-75 ng/ml de sang) dont le taux reste élevé pendant 2 à 3 heures. Spontanément, l'ampleur de la décharge quotidienne de prolactine décroît à partir du 25e jour d'allaitement.
Aspects qualitatifs et quantitatifs de la production laitière
Par rapport au lait de vache, de chèvre ou de brebis, celui de la lapine est beaucoup plus concentré, à l'exception du lactose (tableau 14). Pour le minéraux, on doit souligner la très grande richesse relative et absolue en calcium et en phosphore. A partie de la 4e semaine de lactation, le lait s'enrichit sensiblement en protéines et surtout en lipides (figure 44). Par contre, sa teneur en lactose, déjà faible, diminue encore pour devenir quasi nulle au delà du 30e jour de lactation.
Les teneurs en calcium et en phosphore du lait tendent à s'accroître tout au long de la lactation Celles du potassium et du sodium évoluent en symétrie tendant à maintenir une somme Na + K = constante. Pour les autres minéraux, les teneurs moyennes sont de 30 à 50 ppm pour le zinc, 2 à 4 ppm pour le fer, 1 à 2 ppm pour le cuivre et 0,1 à 0,3 ppm pour le manganèse.
Les matières grasses du lait sont composées principalement de triglycérides, mais contiennent de faibles quantités d'acides gras libres, de phospholipides et de cholestérol. La teneur en acides gras du lait varie en fonction de la nature du régime alimentaire et de l'origine des animaux. Quelques exemples de composition en acides gras sont fournis au tableau 15. Il convient d'en retenir la forte proportion d'acides gras à chaîne courte (C8:0 - C10:0) dont le total peut dépasser la moitié des acides gras totaux, ainsi que la proportion élevée d'acides gras insaturés à 18 atomes de carbone (C18:1 - C18:2 et C18:3). Les teneurs en C8:0 et C10:0 augmentent tout au long de la lactation aux dépens de acides gras à chaîne longue en C16 et en C18, y compris pour les lactations prolongées jusqu'à la 6e ou la 7e semaine après la mise bas.
La production quotidienne de lait croît de 30-50 g les deux premiers jours à 200-250 g vers la fin de la 3e semaine de lactation, voire 300 g/jour pour les souches les plus laitières. Elle décroît ensuite rapidement. La décroissance est plus rapide si la lapine a été fécondée immédiatement après la mise bas
Si la lapine a été fécondée 10 jours après sa mise bas, une décroissance rapide de la production laitière apparaît à compter du 30e jour de lactation. En fait, quelque soit le stade de fécondation par rapport à la mise bas, la production laitière d'une lapine gestante-allaitante se ralentit fortement à compter du 20e jour de gestation et devient nulle au 28-29e jour.
Il existe des différences de forme de courbe de lactation entre individus, notamment en ce qui concerne la persistance. A production totale identique, une croissance rapide avant 18-20 jours et une décroissance rapide ensuite est favorable au taux de survie des lapereaux.
La mesure du poids des lapereaux à 21 jours donne une assez bonne estimation de la lactation totale, car la production laitière 0-21 jours est en étroite corrélation (r = + 0,92) avec la production totale de lait.
Enfin, et c'est un point important, la production laitière de la lapine augmente avec l'effectif de la portée, mais chaque lapereau consomme alors individuellement un peu moins de lait. Toutefois, en fonction du type génétique, l'accroissement de la production avec la taille de la portée cesse au-delà de 10 à 12 lapereaux allaités, voire moins pour les populations non sélectionnées.
Reproduction et environnement
Des lapines éclairées seulement 8 heures sur 24 acceptent beaucoup plus difficilement de s'accoupler que si elles sont soumises à 16 heures d'éclairement chaque jour. Par exemple, des lapines éclairée 8h/24 ont un taux de gestation de 74% quand celui des lapines soumises à un éclairement de 16h/24 est de 84,5%. Un éclairement 12 heures sur 24, permet d'obtenir un résultat intermédiaire.
Effets de l'éclairement
Dans la pratique des élevages rationnels, les locaux de reproduction sont éclairés 15 à 16 heures sur 24, mâles et femelles étant réunis dans la même salle d'élevage si la reproduction se fait en saillie naturelle.
Par ailleurs, comparativement au cas de lapines éclairées régulièrement 16h/24, chez des lapines éclairées 8 heures par jour, le passage brutal à 16 heures d'éclairement par 24 heures une semaine avant la présentation au mâle ou l'insémination permet de faire passer de 54,3% à 71,4% le taux de lapines réceptives conduites selon un rythme 35 jours (intervalle de 35 jours entre deux tentatives de saillies ou 2 inséminations). Enfin, Arveux et Troislouches ont montré en 1994, toujours par rapport à un éclairement continu de 16h/24, que la division des 24 heures en 2 sous-unités de "8 heures d'éclairement + 4 heures d'obscurité" permet d'améliorer la productivité des femelles. En particulier, cela permet de réduire la fonte du cheptel (43% vs 71%), de réduire l'intervalle mise bas - saillie fécondante (19 vs 24 jours), d'obtenir une meilleure fertilité (83% vs 68%) et d'accroître de 4 le nombre de lapereaux sevrés par mère et par an (59 vs 53).
Effets de la température
Des lapines futures reproductrices élevées en ambiance chaude (31°C) ont une croissance nettement ralentie (poids vif réduit de 17%) par rapport à leurs sœurs élevées dans des conditions plus tempérée de 23°C . Cette réduction de croissance est associée à une diminution de 2 du nombre d'ovules pondu à la suite d'un test réalisé à 116 jours.
Toutefois, les réductions de prolificité attribuées aux lapines élevées en ambiance chaude (30-32°C) et illustrées par l'exemple précédant seraient imputables moins à la température elle-même qu'à la réduction du poids corporel entraînée par la baisse du niveau d'ingestion liée à la température élevée. Par contre, il semble que la mortalité embryonnaire augmente lorsque la température dépasse 30-33°C mais, là encore, la part de la réduction d'ingestion n'a pas été faite.
Effets de la saison
La saison est généralement analysée principalement en fonction de la combinaison des effets d'éclairement et de température. Chez le lapin sauvage européen (le lapin de garenne), la reproduction est fortement marquée par la saison.
Les femelles sont en phase de reproduction depuis la fin de l'hiver jusqu'au début de l'été. La période de reproduction peut être allongée ou raccourcie par l'une des deux extrémités en fonction de la température d'une part, mais aussi des disponibilités alimentaires d'autre part.
Un éclairement des lapines domestiques 16 heures sur 24 heures atténue considérablement la variation saisonnière, pour la rendre quasi nulle. Toutefois, quelques difficultés de reproduction peuvent apparaître en fin d'été certaines années, sans relation directe avec la température.
Une étude statistique conduite sur les résultats mensuels de fertilité observés en moyenne sur 4 années, montre qu'il n'y a aucune variation de la moyenne d'un mois sur l'autre seulement une plus forte variabilité en Mai et en Octobre
Une analyse de la prolificité moyenne par trimestre observée par les mêmes auteurs que l'étude précédente, montre également que dans des élevages commerciaux dont les lapines sont éclairées 15-16h/24, il n'est pas possible de trouver un effet répétable de la saison sur 5 années consécutives.
On peut donc considérer que dans les conditions d'un élevage rationnel (commercial), une alimentation de qualité combinée avec un éclairage contrôlé 15 à 16 h/24 heures et un température minimale assurée durant l'hiver (15 à 17°C en général) suppriment les effets saisonniers moyens. Toutefois cela ne veut pas dire que dans un élevage donné, il ne sera pas observé de fluctuation de la fertilité ou de la prolificité d'un mois à l'autre ou d'un trimestre à l'autre.
| Nos expositions |
| Une exposition en bref |
| Marché paysan |
| Foire agricole de Soignies 2004 |
| Foire agricole de Soignies 2005 |
| Le souper annuel |
| Notre site |
| Présentation du club |
| Les canards |
| Les dindons |
| Les grandes volailles |
| Les volailles naines |
| Les lapins |
| Les pigeons |
| Les anatidés |
| Le canard domestique |
| Le canard sauvage "colvert" |
| Les canards de races belges |
| Liste |
| Histoire des dindons |
| Les dindons à Ronquières |
| Liste |
| Anatomie |
| Généralités |
| Liste des grandes volailles |
| L'aile |
| Bréchet |
| Pattes |
| Plumage |
| Queue |
| Squelette |
| Système circulatoire |
| Système digestif |
| Systeme musculaire |
| Système nerveux |
| Système respiratoire |
| Système uro génital |
| La tête |
| Origine |
| Définition |
| Domestication |
| Formation des races |
| Historique |
| Généralités |
| Liste des volailles naines |
| Photos de volailles naines |
| Barbus d'Anvers |
| Barbus d'Uccle |
| Brabançonne |
| Braekel |
| Fauve de hesbaye |
| Herve |
| Mehaigne |
| Tournaisis |
| Watermael |
| Généralités |
| Biologie du lapin |
| Liste des lapins |
| Photos de lapins |
| Argenté Belge |
| Beveren |
| Blanc de Termonde |
| Bleu de Ham |
| Gris perle de Hal |
| Steenkonijn |
| Generalites |
| Liste des pigeons |
| Photos de pigeons |
| Barbet Liegeois |
| Carneau |
| Cravaté gantois |
| Cravaté Liégeois |
| Culbutant belge |
| Haut volant belge |
| Ringslager belge |
| Smerle des flandres |
| Smijter |
| Speelderke |
| Voyageur belge |
| Voyageur Liégeois |
| L'agriculture |
| La fécondation |
| L'origine des oiseaux |
| En péril |
| La domestication |
| Le règne animal |
| Les fermes |
| Evolution |
| Système zoologique |
| Nomenclature |
| Distribution |
| L'adaptation |
| Communication |
| L'habitat |
| La vie sociale |
| Les expressions |
| Un jour un dicton |
| Contes et légendes |
| Fables de La Fontaine |
| A poil ! |
| Un canard boiteux |
| Du coq à l'âne |
| Poser un lapin |
| Fier comme un pou |
| Entre chien et loup |
| Sa langue au chat |
| Connu comme le loup blanc |
| C'est chouette ! |
| Janvier |
| Février |
| Mars |
| Avril |
| Mai |
| Juin |
| Juillet |
| Aout |
| Septembre |
| Octobre |
| Novembre |
| Décembre |